

por Ruben Heleno
Nenhum organismo vivo consegue viver isolado dos organismos que o rodeiam. Pelo contrário, todos os seres vivos estão permanentemente ligados através de uma gigante e complexa rede de interações biológicas. De uma maneira muito simplificada, as plantas (produtores) fixam a energia do Sol e disponibilizam-na em forma de alimento para os animais herbívoros (os consumidores primários), e estes por sua vez aos seus predadores (consumidores secundários). Desde a antiguidade que a importância destas cadeias alimentares para a manutenção das comunidades biológicas é reconhecida. No entanto, mais recentemente surgiu a consciência de que existem vários outros tipos de interações, para além de “quem-come-quem”, que aumentam a complexidade dos sistemas biológicos e que são igualmente vitais para o funcionamento dos ecossistemas. Estas interações incluem o parasitismo, comensalismo, facilitação ou mutualismo, no qual ambas as espécies saem beneficiadas com a interação. Três exemplos clássicos de associações mutualistas são a polinização de flores por animais que em troca recebem uma refeição (tipicamente néctar ou pólen), as associações micorrízicas entre plantas e fungos, e a dispersão de sementes por animais. Neste último caso os animais frugívoros, principalmente aves, mamíferos e répteis, ao alimentarem-se de frutos transportam as suas sementes para novos locais longe da planta-mãe. Desta forma, aumentam as probabilidades de sucesso das sementes, que escapam à competição e aos inimigos naturais da planta-mãe e de sementes conspecíficas, são muitas vezes depositadas em microambientes mais favoráveis à germinação e aumentam a área de distribuição da espécie.
A elevada competição por espaço, nutrientes ou luz, faz com que todas as plantas incapazes de ombrear com as plantas mais competitivas e de manter um lugar em comunidades maduras (ou climax) estejam “condenadas” a ter que aproveitar rapidamente recursos provisórios, criados por perturbações como o fogo, inundações, quedas de árvores ou deslizamentos de terra. Estas plantas são, neste sentido, espécies “fugitivas” que dependem da dispersão das suas sementes para subsistir a longo prazo num intenso “jogo-das-cadeiras” do mundo vegetal. Estima-se que nos trópicos 90% das espécies sejam dispersadas por animais, o que se torna claramente uma vantagem quando é essencial “chegar primeiro”.
Além de ser importante para a dinâmica das comunidades vegetais, a dispersão de sementes é absolutamente crucial para a colonização de ilhas por plantas, especialmente de ilhas oceânicas – que emergiram do fundo do mar, completamente estéreis como é o caso das ilhas vulcânicas. Muitas ilhas vulcânicas apresentam-se aos seus visitantes como um ponto de verdura luxuriante contrastando com os milhares de quilómetros de azul que as separam dos continentes. Para que estas plantas conseguissem cruzar o oceano, as suas sementes (ou outro tipo de diásporo, mas quase sempre as suas sementes) tiveram que flutuar nas longas correntes oceânicas (talassocoria), apanhar boleia interna- (endozoocoria) ou externamente (epizoocoria) de aves e morcegos, ou ser arrastadas por correntes atmosféricas (anemocoria). Uma vez que a chegada de uma nova espécie a uma ilha é um fenómeno incrivelmente raro (algumas estimativas apontam para 1 nova espécie a cada 40.000 anos), a forma mais efetiva de tentar avaliar a eficácia destes diferentes tipos de transporte é comparar a frequência de adaptações para cada um destes tipos de dispersão entre as floras insulares e as floras continentais que lhes deram origem.

Fig.1 O papel dos dispersores de sementes para a regeneração das comunidades vegetais em ilhas e continentes. Esquerda: Uma tutinegra (Pycnonotus tricolor), uma ave tipicamente frugívora, a dispersar as sementes de Sterculia appendiculata [mkunya] no Parque Nacional da Gorongosa em Moçambique. Direita: um papa-moscas-das-Galápagos (Myiarchus magnirostris), tipicamente insectívoro, a dispersar um fruto de unha-de-gato (Zanthoxylum fagara) na Ilha de Santa Cruz – Galápagos. Em baixo: sementes da árvore tóxica de manzanillo (Hippomane mancinela) a crescer num excremento de tartaruga gigante (Chelonoidis nigra), na ilha Española – Galápagos, Ruben Heleno©.
Os poucos estudos disponíveis sugerem uma destacada relevância para o transporte por correntes oceânicas, com um efeito menor do transporte interno por animais, e um efeito quase nulo ou mesmo dificultador da colonização para sementes adaptadas ao transporte externo por animais ou por correntes atmosféricas (Fig. 1).
Curiosamente, a barreira que separa as ilhas dos continentes também faz com que haja menos dispersores disponíveis para as plantas que conseguirem fixar-se nas ilhas. Este vazio ecológico criado por um filtro imposto a espécies pouco dispersivas, torna os sistemas insulares muito característicos, tendo geralmente baixa biodiversidade, alto grau de endemismos e alta densidade de indivíduos que muitas vezes ocupam nichos ecológicos inesperados. Entre alguns destes exemplos encontram-se o importante papel de répteis como dispersores de sementes em ilhas, ou de aves granívoras e insectívoras como consumidores de néctar e polinizadores de flores.
Enquanto alguns tipos de interações biológicas, como por exemplo a relação parasita-hospedeiro, são altamente especializadas, as relações mutualistas e principalmente a dispersão de sementes são geralmente generalistas. Normalmente as redes de dispersão caracterizam-se por um forte “aninhamento”, em que a esmagadora maioria das espécies (incluindo as mais especialistas) interage com um “núcleo duro” de espécies altamente generalistas. Este elevado grau de generalização é uma espada de dois gumes. Por um lado torna as redes de dispersão resistentes à extinção, global ou local, de espécies, uma vez que outras espécies podem assumir a função das espécies extintas (redundância de dispersores).
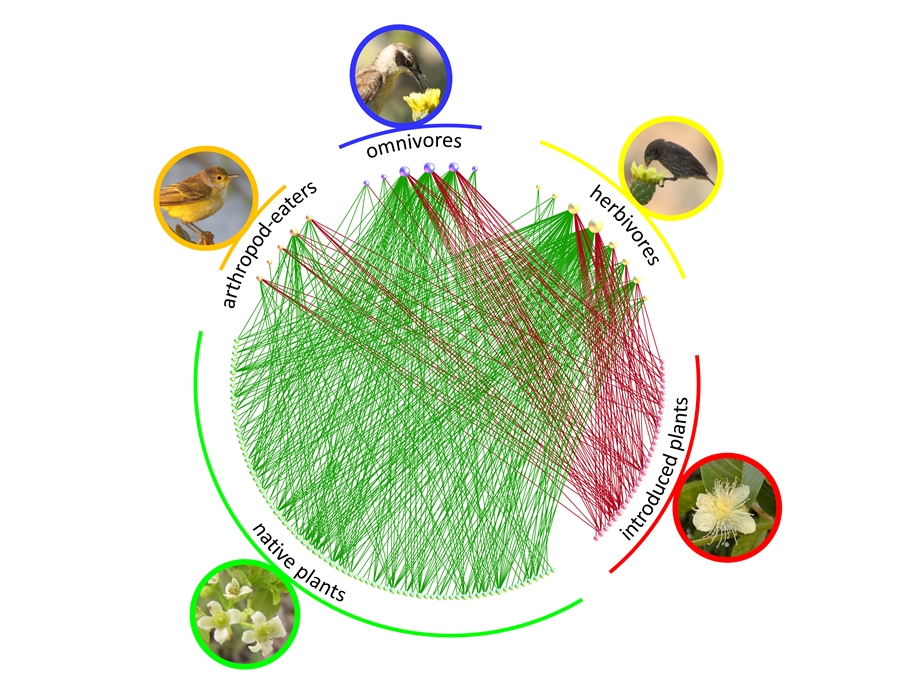
Fig. 2 Rede de transporte de pólen por aves nas Galápagos. As 19 espécies de aves amostradas, incluindo três grupos funcionais, visitam as flores e transportam pólen de 106 espécies de plantas incluindo uma elevada percentagem de plantas introduzidas (Traveset el al. in press).
Por outro lado, também torna estas redes muito vulneráveis à chegada de espécies exóticas, nomeadamente de espécies invasoras que são muito rapidamente integradas na dieta de animais dispersores e/ou polinizadores e que frequentemente facilitam a invasão (e.g. Fig. 2).
Não é por isso surpresa que a maioria das plantas vasculares invasoras produzam frutos carnosos ou outras estruturas para atrair dispersores. Este “serviço” de dispersão prestado às plantas introduzidas, normalmente faz-se às custas de uma perda de qualidade do serviço prestado à comunidade de plantas nativas. Esta “troca” pode alterar profundamente a estrutura das redes de dispersão de sementes e consequentemente a dinâmica da vegetação e dos ecossistemas, por regra conduzindo a uma perda de diversidade. Outras vezes é a chegada de um novo dispersor introduzido ou a perda de um dispersor nativo que altera a quantidade e a qualidade do serviço de dispersão, introduzindo novos filtros para o recrutamento de plantas. A peculiaridade dos ecossistemas insulares, com uma história evolutiva em isolamento, torna-os mais vulneráveis à chegada de espécies altamente competitivas. Outro grupo de risco, no que se refere a dispersão de sementes, são as plantas com frutos e sementes grandes, uma vez que os seus dispersores (também eles grandes) são tendencialmente mais afetados pela sobre-exploração, perda e fragmentação dos habitats.
Infelizmente, muitas destas alterações, como a fragmentação e perda de habitat, invasões biológicas, poluição e alterações climáticas, são globais e têm efeitos muitas vezes sinergísticos e difíceis de prever. As redes ecológicas têm-se mostrado uma ferramenta muito útil para prever alterações no funcionamento e na dinâmica dos ecossistemas, bem como para planear e monitorar a sua conservação recuperação. Por exemplo, está demonstrado que o generalismo típico das redes de dispersão de sementes facilita os programas de restauração de habitats que se dedicam a restaurar funções vitais dos ecossistemas e pode ainda permitir o uso de “substitutos funcionais” para substituir ou reforçar as funções em declínio devido à extinção de espécies mais sensíveis. A aplicação da “teoria de redes” em ecologia é por isso particularmente útil para estudar o funcionamento dos ecossistemas, oferecendo um enquadramento no qual se podem testar hipóteses relativas a importantes problemas ecológicos, como a perda de biodiversidade. Este enquadramento permite avaliar simultaneamente a composição (i.e. as espécies), a estrutura (i.e. as interações) e o funcionamento (i.e. os serviços ecológicos prestados), considerando a totalidade dos ecossistemas mas sem perder de vista as espécies responsáveis por esses mesmos serviços.
Infelizmente as visionárias palavras de Charles Elton há quase 70 anos são cada vez mais atuais:
We must make no mistake: we are seeing one of the great historical convulsions in the world’s fauna and flora. (…) Unless one merely thinks man was intended to be an all-conquering and sterilizing power in the world, there must be some general basis for understanding what is the best to do. This means looking for some wise principle of co-existence between man and nature, even if it has to be a modified kind of man and a modified kind of nature.
Charles Elton, 1958 in The Ecology of Invasions by Animals and Plants.
Referências usadas:
Elton, C. S. (1958) The ecology of invasions by animals and plants, edn. Methuen, London.
Heleno, R., Olesen, J. M., Nogales, M., Vargas, P. & Traveset, A. (2013a) Seed-dispersal networks in the Galápagos and the consequences of plant invasions. Proceedings of the Royal Society B-Biological Sciences, 280, 20122112.
Heleno, R., Ramos, J. & Memmott, J. (2013b) Integration of exotic seeds into an Azorean seed dispersal network. Biological Invasions, 15, 1143-1154.
Heleno, R. H., Ceia, R. S., Ramos, J. A. & Memmott, J. (2009) The effect of alien plants on insect abundance and biomass: a food web approach. Conservation Biology, 23, 410-419.
Heleno, R. H. & Vargas, P. (in press) How do islands become green? Global Ecology and Biogeography.
Nogales, M., Heleno, R., Traveset, A. & Vargas, P. (2012) Evidence for overlooked mechanisms of long-distance seed dispersal to and between oceanic islands. New Phytologist, 194, 313-317.
Traveset, A., Heleno, R. H. & Nogales, M. (2014) The ecology of seed dispersal. Seeds: The ecology of regeneration in plant communities (ed. by R.S. Gallagher), pp 62-93. CABI, Oxfordshire, UK.
Traveset, A., Olesen, J. M., Nogales, M., Vargas, P., Jaramillo, P., Antolín, E., Trigo, M. M. & Heleno, R. (in press) Bird-flower visitation networks in the Galápagos unveil a widespread interaction release. Nature Communications.
Traveset, A. & Richardson, D. M. (2006) Biological invasions as disruptors of plant reproductive mutualisms. Trends in Ecology & Evolution, 21, 208-216.
Vargas, P., Heleno, R., Traveset, A. & Nogales, M. (2012) Colonization of the Galápagos Islands by plants with no specific syndromes for long-distance dispersal: a new perspective. Ecography, 35, 33-43.



Comentários